Будет рассматриваться на примере --- ольно крупные размеры. К примеру, взрослый малый рабочий достигает 2,5 метров в длину (с вытянутыми нижними конечност ---трех четко выраженных отделов.
Верхний отдел (лофофор) представляет собой несегментированную хитиновую трубку, увенчанную шестью перистыми придатками - антеннулами --- хеморецепторов, в том числе так называемых овоидных купул - рецепторов феромонов. В верхнем отделе лофофора расположены также два простых глазка, состоящих из покровной линзы, внутренней стекловидной массы и сенсорной пластинки. По бокам лофофора расположены верхние конечности, каждая из которых имеет два отдела: мускульную трубку и щупальцевый венчик из 10 - 15 щупалец. Эндоскелет у верхних конечностей отсутствует, экзоскелетные образования представлены эллиптическим сочленовным элементом, расположенным у места прикрепления конечности с лофофором, и приблизительно сотней верхних и нижних черепицеобразных пластин разного размера и формы, покрывающих мускульную трубку и каждое из щупалец. Черепицеобразные пластины соединяются пленчатыми связками из более тонкого хитина. Благодаря такому устройству каждая точка верхней конечности обладает шестью степенями свободы. Для фиксации конечности в определенном положении используются сочленовные зубчики, расположенные на дистальных поверхностях черепицеобразных пластинок. Замыкание сочленовных зубчиков осуществляется особыми мышцами (mm. Fixatores limbi superi --- доскелетной основой лофофора является копьевидный отросток хордального кольца (см. ниже), проходящий через соединительное отверстие в бранхиофор. К копьевидному отростку подвешены антеннальная артерия, ветви которой питают верхние конечности, антеннулы и простые глазки, антеннальная вена, ответственная за венозный отток из указанных областей, мощное копьевидное нервное сплетение с расположенными спирально вокруг копьевидного отростка девятнадцатью-двадцатью двумя нервными ганглиями I порядка (спиральными ганглиями), ответственное за чувствительную и двигательную иннервацию верхних конечностей, легких и псевдосиринкса (см. ниже), а также антеннулярные и глазковые нервы.
Средний отдел (бранхиофор) имеет вид четырех хитиновых пузырей (тек), соединенных у основания. На верхушке каждой из тек имеются отверстия, прикрытые фильтрационными складками. Меньшие, верхние пузыри - сиринготеки - содержат парный орган вокализации - псевдосиринкс. Большие, нижние пузыри - бранхиотеки - содержат парные "легкие" чужих. На передней поверхности бранхиофора в месте слияния оснований тек (decussatio tнecarum) находится ротовое отверстие, окруженное хватательно-пожирательным органом. Последний (у малого рабочего) состоит из четырех хитиновых крючьев длиной около 25 см, соединенных общей фиброзно-мышечной мембраной. От ротового отверстия начинается глотка, идущая книзу и кзади и заканчиваясь в области верхнего сфинктера желудка. Позади глотки расположено основание копьевидного отростка, продолжающегося кверху в лофофор, и связанные с ним сосуды и нервы (см. выше).
Нижний отдел (висцеротека) представляет собой несегментированный эллипсоид из плотного хитина. В области верхнего полюса висцеротеки находится хордальное кольцо - основа эндоскелета чужих. Это бубликовидное образование, построенное из оссеоидной ткани. По нижней поверхности хордального кольца прикрепляется общая брыжейка - фиброзная пленка, выстланная мезотелием. Общая брыжейка состоит из 5 отделов: 1) брыжейка желудка; 2) брыжейка сердечных колец; 3) брыжейка зеленой железы; 4) брыжейка тонкой кишки; 5) брыжейка толстой кишки.
В толще общей брыжейки заложены основные кровеносные сосуды - парахордальное (нижнее) и бранхиальное (верхнее) кольца. От внутренней поверхности хордального кольца отходят гиалиновые пластины, образующие капсулу мозга. Органы, подвешенные на брыжейке, заполняют все внутреннее пространство висцеротеки. В области ее нижнего полюса находится анальное отверстие, куда открываются толстая кишка и проток зеленой железы Верхние конечности, расположенные по бокам висцеротеки, не имеют эндоскелета. Миоэпителиальная основа нижних конечностей состоит из двух отделов - мускульной трубки и терминального расширения. Экзоскелет нижней конечности, в отличие от верхней, представлен эллиптическим сочленовым элементом, двумя трубчатыми образованиями - бедром и голенью, соединенными двумя переходными пластинками. Плотную кутикулу терминального расширения пронизывают тяжи, оканчивающиеся мощными опорными шпор эндоскелет чужих состоит из двух основных образований - трехгранного хордального кольца и отходящего вертикально от верхней поверхности последнего копьевидного отростка. Внутренняя поверхность хордального кольца продолжается в три тонкие треугольные гиалиновые пластинки. Капсула мозга является дупликатурой гиалиновых пластинок. В месте отхождения копьевидного отростка от хордального кольца разделены треугольной щелью, через которую проходят антеннулярный, глазковый, общий висцеральный нервы, а также корешки копьевидного и педального сплетений. ---построен из оссеоидной ткани - микроскопических кристаллов арабонита и карбоната железа, спаянных бесструктурным --- сульфатированных полисахаридов и фибриллярного кератиноподобного белка. Клеточные элементы - хордобласты - находятся только в тонком (100-200 мкм) поверхностном слое. В гиалиновых пластинках минеральные спикулы --- Покровы тела чужих представлены миоэпителиальной тканью I рода. Экзоскелетные структуры являются продуктом секреции миоэпителия. Все они пронизаны тонкими каналами, содержащими цитоплазматические отростки миоэпителиоцитов. Базальные сократительные волокна миоэпителиоцитов сливаются вместе, образуя мышечную основу конечностей и общий субкутикулярный мускульный пласт, имеющийся во всех отделах тела, но особенно развитый в нижней части лофофора. Изнутри субкутикулярный мышечный пласт выстлан мезотелием. В висцеротеке между субкутикулярным пластом и мезотелием залегает дольчатое скопление жировых клеток (жировая подушка).
Органы дыхания и вокализации чужих представлены парным псевдосиринксом (органом вокализации), находящимся в двух верхних теках бранхиофора, и парными "легкими" в двух нижних теках. Как псевдосиринкс, так и "легкие" являются миоэпителиальными органами, образованными сетью трабекул. Внутри каждой трабекулы находятся сосуды и сократительные волокна. Снаружи трабекулы покрыты миоэпителием IV рода. Трабекулы "легких " имеют диаметр около 50 мкм, при этом толщина аэрогемального барьера составляет 15 мкм. Трабекулы псевдосиринкса знасительно толще (от 500 мкм до 7 мм) и имеют выраженный каркас из эластических волокон. Кроме того, часть из них снабжена хитиновыми гребнями и крючками. Каждый миоэпителиальный орган имеет корень, находящийся у основания соответствующей теки. В области корня проходят магистральные сосуды - ветви брнхиального кольца - и начинается дренажный проток, открывающийся в глотку у основания хватательнопожирательного органа. Дренажный проток представляет собой трубку длиной около 30 мм, образованную миоэпителием IV рода.
Органы кровообращения.
Кровеносная система чужих - замкнутого типа. Основными ее элементами являются два крупных кольцевых сосуда - парахордальное и бранхиальное кольца - заложенные в толще общей брыжейки, два соединительных протока и заложенные в их стенках пульсирующие мышечные подушки - --- ижнее, парахордальное, кольцо состоит из артериального и венозного сегментов. Диаметр венозного сегмента меньше артериального. От артериального сегмента отходят артерии, питающие глотку, желудок, кишечник, зеленую железу, у высших особей - гонады. В венозный сегмент впадают соответствующие вены.
Верхнее, бранхиальное, кольцо состоит из двух сегментов - бранхиального (от места впадения венозного соединительного протока до места впадения легочных вен) и антеннального (от места впадения легочных вен до места отхождения артериального соединительного протока). В бранхиальный сегмент, по которому течет неоксигенированная кровь, впадают церебральная, сирингоидная и антеннальная вены, далее от него отходят две легочные артерии (по одной к каждому легкому). В антеннальный сегмент впадают две легочные вены, несущие оксигенированную кровь от легких. От него отходят церебральная артерия (к мозгу), сирингоидная артерия (к псевдосиринксу) и антеннальная артерия (к органам лофофора). Диаметр бранхиального кольца суживается в местах впадения вен и расширяется в местах отхождения артерий.
Венозный соединительный проток соединяет венозный сегмент парахордального кольца и бранхиальный сегмент бранхиального кольца.
Артериальный соединительный проток соединяет антеннальный сегмент бранхиального кольца и артериальный сегмент парахордального кольца.
Каждый проток окружен "муфтой" из циркулярно расположенных гладких миоцитов. Соединительные протоки и их мышечные "муфты" вместе составляю ---в участках парахордального и бранхиального колец, находящихся между устьями протоков (межпротоковый, или третий, сегмент) в стенках сосудов также заложены циркулярные пласты гладких миоцитов. Их функция - регуляция степени смешения артериальной и венозной крови.
Органы пищеварения чужих представлены глоткой, мышечным и железистым желудком, тонким и толстым кишечником. Все указанные образования сформированы миоэпителиальной тканью II и III родов. Мышечный компонент миоэпителия формирует функциональный синцитий с перекрестным ходом сократительных волокон, ответственный за перистальтику.
Глотка сформирована миоэпителиальной тканью II рода и представляет собой трубчатое образование длиной около 25 см, открывающееся в мышечный желудок в области его верхнего сфинктера. В верхний отдел глотки, тотчас позади хватательно-пожирательного органа открываются дренажные протоки легких и псевдосиринкса, по два с каждой стороны.
Желудок сформирован миоэпителиальной тканью II рода и представляет из себя мешковидное образования емкостью до 20 литров. В области впадения глотки и отхождения тонкой кишки мышечный компонент миоэпителия образует два циркулярных утолщения - верхний и нижний сфинктеры.
Тонкая кишка сформирована миоэпителиальной тканью III рода и представляет из себя трубку длиной около 1 м. Внутренняя поверхность тонкой кишки покрыта складками, имеющими спиральный ход и весьма напоминает таковую у земных селахий. Кнаружи от мышечного синцития располагаются многочисленные заключенные в соединительнотканную капсулу железки, продуцирующие смесь гидролаз. Их выводные протоки открываются на внутренней поверхности кишки.
Толстая кишка сформирована миоэпителиальной тканью II рода и представляет собой трубку с относительно гладкой внутренней поверхностью длиной около 10 м. Толстая кишка открывается наружу анальным отверстием. Вокруг последнего мышечный компонент миоэпителия формирует циркулярное утолщение - сфинктер. Ниже нижнего края сфинктера отдельным отверстием открывается проток зеленой железы.
Орган детоксикации и поддержания солевого гомеостаза - зеленая железа - представляет собой бесформенное дольчатое образование зеленого цвета массой около 3 кг, сформированное миоэпителиальной тканью IV рода. Секрет зеленой железы - высокомолекулярный полисахарид, обладающий свойствами универсального сорбента для катионов, анионов и полярных органических веществ с одним или несколькими ароматическими ядрами. С этим секретом выделяется как избыток солей, так и неметаболизируемые органические токсины.
Нервная система чужих состоит из центрального отдела (мозг) и периферического отдела (антеннулярный, глазковый, общий висцеральный нервы, копьевидное, спиральное сплетение и т. д.).
Мозг чужих, как указывалось выше, находится в капсуле, образованной расщеплением гиалиновых пластинок. Треугольная щель капсулы затянута соединительнотканной мембраной. Капсула зполнена жидкостью, по плотности близкой к веществу мозга.
Макроскопически мозг чужих имеет пять хорошо выраженных отделов.
1. Висцеральный мозг - шаровидное образование с гладкой поверхностью, находящееся в задненижней части капсулы мозга. От верхней поверхности висцерального мозга отходит непарный общий висцеральный нерв. Боковые поверхности соединены со стеблем соматического мозга посредством двух коннектив. На разрезе висцеральный мозг состоит из:
· наружного слоя серого вещества толщиной около 1,5 мм (образован телами нейроцитов); · слоя белого вещества толщиной около 5 см, образованного отростками нейроцитов; · центрального серого ядра, образованного телами нейроцитов и состоящего из нескольких трудно различимых клеточных групп.
Функцией висцерального мозга является чувствительная и двигательная иннервация внутренних органов, а также контроль метаболизма, эндокринных функций и секреции феромонов, осуществляемый, в основном, нейросекреторными элементами центрального серого ядра и иннервирующими экзо- и эндокринные железы волокнами общего висцералльного нерва.
2. Соматический мозг - образование неправильной формы, имеющее вид цилиндра с тремя утолщениями. - щупальцевым, сирингобранхиальным и педальным. Соматический мозг находится в задневерхней части капсулы мозга. От стебля соматического мозга в его заднем отделе (между сирингобранхиальным и педальным утолщениями) отходят две коннективы к висцеральному мозгу, а в среднем отделе (между сирингобранхиальным и щупальцевым утолщениями) - две коннективы к сенсорному мозгу. Передний отдел стебля, равположенный кпереди от щупальцевого утолщения, переходит в центральный волокнистый пучок межуточного мозга. На боковых поверхностях утолщений имеются переходные борозды, в которых расположены корешки и стволики сегментарных нервов. Каждый сегментарный нерв образуется путем слияния верхнего (чувствительного) и нижнего (двигательного) корешков. От щупальцевого утолщения отходит по 2, от сирингобранхиального - по 5 и от педального - по 3 сегментарных нерва с каждой стороны. Сегментарные нервы покидают капсулу через перфорирующие ходы на ее нижней поверхности. 2 сегментарных щупальцевых и 5 сегментарных сирингобранхиальных нервов огибают капсулу мозга и парахордальное кольцо снизу вверх и спереди назад, проходя через отверстия в общей брыжейке, и направляются к основанию копьевидного отростка, где формируют копьевидное сплетение. Три педальных нерва направляются к нижневнутренней поверхности висцеротеки, где формируют педальное сплетение. На разрезе соматический мозг состоит из периферического слоя серого вещества толщиной около 2 мм и образованной белым веществом сердцевины. В области утолщений толщина периферического слоя достигает 4 мм, переходная борозда разделяет его на две четко различимые пластинки - большую (сенсорную и ассоциативную) и меньшую нижнюю (моторную).
3. Сенсорный мозг - парное образование, расположенное в передневерхней части капсулы мозга. Состоит из двух долей (зрительной и антеннулярной), разделенных пограничной бороздой. От верхней поверхности долей отходят глазковый и три антеннулярных нерва. Половины сенсорного мозга соединены валиком - широким плоским пучком нервных волокон. От нижней поверхности валика отходят две передних коннективы - длинных тонких пучка волокон, отходящих вниз к конечному мозгу, две средних коннективы, идущих к межуточному мозгу, и две задних коннективы, направляющихся назад к среднему отделу стебля соматического мозга.
4. Межуточный мозг представляет собой продолжение стебля соматического мозга и занимает центральную часть капсулы мозга. Основу межуточного мозга составляет центральный волокнистый пучок - мощный пласт нервных волокон, между которыми заложено множество (до пятидесяти) клеточных скоплений - межуточных ядер, образующих сенсорную часть церебрального магнитосенсорного аппарата. Снаружи центральный волокнистый пучок покрыт слоем серого вещества слоем толщиной до 3 мм - продолжением серого вещества стебля соматического мозга. Клетки этого слоя ответственны за организацию сложных моторных актов. К боковой поверхности межуточного мозга подходят две коннективы от сенсорного мозга. На задней поверхности межуточного мозга находится полость - магнитосенсорная булла, заполненная бесцветной прозрачной жидкостью - магнитолимфой. Внутри магнитосенсорной буллы находится мышечный генератор магнитосенсорного аппарата. Стенки магнитосенсорной буллы сформированы волокнистой нейроглией, среди которой проходят отдельные нервные волокна, отходящие от центрального нервного пучка и иннервирующие мышечный генератор.
5. Конечный мозг находится в передненижней части капсулы мозга и представляет собой как бы нарост размером 25 - 30 на 15 - 20 см (у разных форм чужих его размеры варьируют) на конце центрального волокнистого пучка межуточного мозга. Внешне конечный мозг более всего напоминает грецкий орех. На разрезе он состоит из пластин серого вещества, разделенных тонкими прослойками проводящих волокон. Конечный мозг ответственен за организацию наиболее сложных форм адаптивного поведения, в том числе речи и интеллектуальной деятельности.
Органы чувств:
Чужие обладают следующими видами рецепторов:
1) дистантные экстерорецепторы, сгруппированные в особые органы чувств - орган зрения (простые глазки у большинства форм рабочих и некоторых видов боевых особей или более-менее развитые фасеточные глаза у прочих), орган хеморецепции и восприятия воздушного давления (антеннулы), церебральный магнитосенсорный аппарат - своеобразный заменитель предметного зрения, использующийся также для записи, воспроизведения и обмена информацией в трехмерных динамичных образах (см. Язык чужих); 2) дистантные экстерорецепторы, не сгруппированные в органы чувств - рассеянные по покровам тела пузырьковые сенсиллы, ответственные за слуховую и вибрационную чувствительность; 3) контактные экстерорецепторы - волосковые сенсиллы и свободные нервные окончания, локализованные на поверхности и в толще экзоскелетных структур, ответственные за восприятие давления, растяжения и температуры внешней среды; 4) соматические интерорецепторы - свободные и инкапсулированные нервные окончания в толще мышечного компонента миоэпителия, образующего покровы тела, легкие, псевдосиринкс и конечности, ответственные за восприятие мышечных сокращений и болевую чувствительность; 5) висцеральные интерорецепторы - свободные и инкапсулированные нервные окончания, локализованные в стенках сосудов и полых органов, а также отростки клеток центрального серого ядра висцерального мозга, ответственные за восприятие концентрации основных метаболитов и газов крови, а также тонуса стенок и моторики сосудов и полых органов.
Орган зрения малого рабочего представляет собой простой глазок, состоящий из: 1) покровной линзы - экзоскелетного образования линзовидной формы, обладающего высокой прозрачностью; 2) стекловидной массы - густой вязкой жидкости, на поверхности которой находится покровная линза; 3) сенсорной пластинки, состоящей из 30-40 светочувствительных нейроэпителиоцитов и расположенных между ними 50-60 секреторных клеток, ответственных за продукцию и резорбцию стекловидной массы. Глазок заключен в соединительнотканную капсулу, основное вещество которой окрашено в темный цвет. Отростки нейроэпителиоцитов прободают капсулу, подстилающий глазок мышечный компонент миоэпителия и идут к копьевидному отростку, собираясь в глазковый нерв.
Хеморецепторный орган - антеннула - является выростом экзоскелета лофофора. Основу антеннулы составляет полый стержень, от которого отходят ветви первого, второго и третьего порядка. На поверхности ветвей третьего порядка (не являющихся выростами экзоскелета, а представляющие собой миоэпителиальные выросты, способные прятаться в полостях ветвей второго порядка) находятся т. Н. Росяные купулы - скопления нейроэпителиальных клеток, покрытые "шапочками" из вязкой слизи. Отростки нейроэпителиальных клеток проходят в полостях ветвей и стержня, сливаясь у его основания в антеннулярный нерв. Основание стержня глубоко вдается в мышечный компонент миоэпителия лофофора, к нему крепятся несколько мышечных пучков радиального направления, осуществляющих движения антеннулы.
Церебральный магнитосенсорный аппарат является основным органом дистантной рецепции малого рабочего. Принцип работы данного органа чувств - регистрация переменного магнитного поля сложной конфигурации, генерируемого самим аппаратом, под влиянием окружающих тел. Основными отделами церебрального магнитосенсорного аппарата являются мышечный генератор, создающий электромагнитный импульс сложной конфигурации, и межуточные сенсорные ядра, воспринимающие возмущение переменного электромагнитного поля под действием окружающих тел. Воссоздание трехмерной картины окружающего происходит в основном в конечном мозге. Мышечный генератор представляет собой совокупность 12-15 рыхлых отростчатых шаровидных образований, взвешенных в магнитолимфе магнитосенсорной буллы. Каждое из этих образований - генерирующих модулей - состоит из более плотного шаровидного тела, окруженного облаком паутинообразных отростков, соединяющих тело с другими. В центре тела находится небольшой удлиненный кристалл карбоната железа. Гистологически генерирующие модули построены из особой возбудимой ткани, сходной одновременно с нервной ткань
--- овой и вибрационный рецептор - пузырьковая сенсилла - представляет собой гиалиновидную полость в экзоскелете, заполненную воздухом. Наружная стенка полости тонка и под воздействием колебаний резонансной частоты приходит в движение. Внутренняя стенка полости состоит из более мягкого хитина, пронизанного нервными окончаниями. Резонансные частоты различны для сенсилл, расположенных на разных участках тела, что обеспечивает восприятие звука и вибрации в широком диапазоне частот. Общее число пузырьковых сенсилл на теле малого рабочего - более 27000.
Дата: Воскресенье, 09.01.2011, 10:58 | Сообщение # 2
Местный житель планеты N
Группа: Администраторы
Сообщений: 8307
Статус: Offline
Анатомия и Физиология "Чужих"
Отсканированная копия оригинала текста, около 10 лет назад появившегося в российских уфологических кругах и распространенного сначала в ФИДО, а затем в Интернете. Автор неизвестен. Нумерация листов условная.
Дата: Воскресенье, 09.01.2011, 11:57 | Сообщение # 5
Местный житель планеты N
Группа: Администраторы
Сообщений: 8307
Статус: Offline
Рецензия на текст "Анатомия чужих"
Э.Н. Золотарева, кандидат биологических наук.
В интернете на нескольких сайтах появился весьма любопытный документ в виде фотокопий с 27 страниц машинописного текста. Этот текст представляет собой, по-видимому, фрагменты предполагаемого доклада, он плохо отредактирован и не является законченным ни по форме, ни по содержанию, кроме того он был частично уничтожен. Тем не менее, ввиду важности содержащейся в нем информации, представляется совершенно необходимым провести его экспертную оценку.
В докладе представлены результаты исследования неизвестных (во всяком случае неизвестных современной официальной науке) форм живых организмов, условно называемых «Чужими».
Эти животные названы так, вероятно, потому, что не принадлежат к человеческому виду, но в то же время являются, якобы, высокоразумными. В уцелевшей части текста нет какой-либо информации относительно их происхождения или истории обнаружения. Имеется только указание на какие-то события, происходившие в 1965 г., по-видимому, начала исследований.
Эти исследования, если они действительно проводились, были выполнены на высоком профессиональном уровне и требовали участия специалистов различных специальностей: физиологов, биохимиков цитологов. С другой стороны, ввиду высокого уровня засекреченности проблемы, круг участников исследования неизбежно должен был быть ограниченным и, по-видимому, не всегда имелась возможность привлечь к работе исследователей, занимающихся соответствующими разделами физиологии беспозвоночных (не говоря уже об исследованиях высшей нервной деятельности, которые определенно должны были бы проводиться специалистами в физиологии человека).
Применяемая терминология в целом свидетельствует о профессионализме авторов доклада. В отдельных случаях употребляются нетрадиционные названия. Это может быть вызвано частично ограничением числа узких специалистов по объективным причинам, обусловленное секретностью. Но, с другой стороны, обнаружение подобной терминологии в тексте, особенно в тех случаях, когда описание явления опережает по времени его официальное открытие (ниже рассматривается случай с гемоцианином) представляет наибольший интерес для нашего обсуждения, поскольку может свидетельствовать о данных, полученных непосредственно из практического опыта, а не из литературы.
Оценить подлинность документа непросто, поскольку отдельные разделы доклада могут быть достаточно хорошо поняты только узкими специалистами, и в особенности, физиологами в энтомологии. К сожалению все обращения за консультативной помощью к энтомологам или специалистам в физиологии насекомых остались без ответа. Поэтому мы попытаемся, во-первых, самостоятельно определить происхождении «Чужих». Во-вторых, мы затронем отдельные вопросы биохимии и физиологии крови, при этом мы намеренно не будем касаться многих аспектов исследования, в т.ч. и физиологии ЦНС.
1). Относительно происхождения «Чужих». Достаточно странно упоминание в начале каждого раздела сравнения с земными позвоночными, поскольку речь в докладе идет о формах, имеющих безусловно земное происхождение. Об этом свидетельствует клеточное строение организмов «Чужих», наличие хромосомного аппарата, их способность дышать кислородом воздуха и т.д. Наличие хитинового экзоскелета и конечностей свидетельствует об их принадлежности к беспозвоночным членистоногим, а описанная физиология и особенности поведения позволяют отнести их к классу насекомых (Insecta). Поэтому остается непонятной фраза о «(ко)лониальных червеподобных организмах, сочетающих признаки земных мшанок». Создается впечатление, что авторы доклада не были сильны в биологии беспозвоночных или же специально уводили вопрос в сторону.
Далее. В тексте многократно упоминаются кастовые особенности «Чужих», как то: малый рабочий, боевые особи, высшие особи. Это позволяет отнести изучаемые формы к высокообщественным насекомым, поскольку имеются морфологически- и физиологически-выраженные кастовые различия между представителями вида. Как известно, эусоциальные (общественные) колонии могут образовывать представители двух отрядов насекомых, а именно: отряда Перепончатокрылых (Hymenoptera), куда относятся осы, пчелы и муравьи, и отряда Термитов (Isoptera). Следует отметить, что эти два отряда имеют далеко не родственное происхождение и независимые пути возникновения эусоциальности.
В докладе есть указание на то, что размножение «Чужих» осуществляется парой высших особей, которые диплоидны (т.е. содержат двойной набор хромосом). Поэтому Перепончатокрылых стоит, по-видимому, исключить из дальнейшего обсуждения, поскольку известно, что только самки этих насекомых диплоидны (и высшие и рабочие), самцы же – гаплоидны (т.е. содержат одинарный набор хромосом); кроме того, самцы недолгоживущи и поэтому Перепончатокрылые не размножаются парами. Таким образом, можно предположить, что «Чужие», вполне вероятно, могут быть родственны современным формам термитов и имели с ними общую предковую форму.
Еще одну параллель между «Чужими» и Термитами можно усмотреть в способе укладки хромосом. Так кариотип «Чужих» «ЯВЛЯЕТСЯ КРАЙНЕ ИЗМЕНЧИВЫМ: В ХРОМОСОМНОМ НАБОРЕ ПОСТОЯННО ПРИСУТСТВУЮТ МЕЛКИЕ ФРАГМЕНТЫ ХРОМОСОМ, КОЛЬЦЕВИДНЫЕ ХРОМОСОМЫ; ЧИСЛО, РАЗМЕР И ХАРАКТЕР ДИФФЕРЕНЦИАЛЬНОГО ОКРАШИВАНИЯ КРУПНЫХ ХРОМОСОМ КРАЙНЕ ВАРИАБЕЛЬНЫ». Явление это связывается авторами доклада с развитием «АЛЛОЦИД-ЗАВИСИМОЙ ТРАНСДУКЦИИ». Употребление этого термина в данном контексте малопонятно, т.к. обычно под генетической трансдукцией понимают перенос генетического материала из одной клетки в другую с помощью, например, умеренного фага. С другой стороны, изменчивость кариотипа является хорошо известным явлением для мужских особей рабочих Термитов. Она заключактся в частичной транслокации различных частей хромосом (в том числе и половых) в процессе мейоза, в образовании фрагментов и кольцевых хромосом [Vincke P.P., Tilquin J.P. A SEX-LINKED RING QUADRIVALENT IN TERMITIDAE (ISOPTERA). Chromosoma , 1978, 67, 151–156; Luykx, P. A CYTOGENETIC SURVEY OF 25 SPECIES OF LOWER TERMITES FROM AUSTRALIA. Genome., 1990, 33, 80–88; Martins, V.G. KARYOTYPE EVOLUTION IN THE TERMITIDAE (ISOPTERA).Sociobiology, 1999, 34, 395–405.]. Как видно, первые публикации на эту тему появились в 1978 г. и могли бы стать источником сведений. Однако, авторы доклада демонстрируют неосведомленность. Сейчас активно обсуждается вопрос о роли этого явления в развитии эусоциальности Термитов [Thorne B.L. EVOLUTION OF EUSOCIALITY IN TERMITES. Annu. Rev. Ecol. Syst. 1997. 28:27–54; Roisin Y. CASTE SEX RATIOS, SEX LINKAGE, AND REPRODUCTIVE STRATEGIES IN TERMITES. Insectes soc.2001, 48, 224–230].
Можно отметить еще немало общих черт у Чужих и Термитов. Отрыгивание пищи, например, низшими рабочими для питания высших и боевых особей, необходимость связи с источником влаги, светобоязнь. Среди особенностей внешнего строения можно отметить существование у разных каст простого и сложного зрения. Сложным зрением обладают, как правило, высшие особи. Кроме того имеются два простых глазка (у представителей перепончатокрылых: ос, пчел и муравьев, в отличие от термитов, имеется 3 простых глазка). Очень сходным является поведение, обусловленное феромонами. Более подробная характеристика этих животных может быть дана энтомологом.
Предполагается, что группа Термиты произошла от архаичных таракановых в триасовом периоде (250-200 млн. лет назад). Нельзя исключить, что в этот период началась эволюция эусоциальных форм насекомых, приведшая, в конечном итоге к «Чужим»). Эта группа животных могла по каким-то причинам подвергнуться длительной изоляции и действию экстремальных факторов. А отрезок в сотни миллионов лет мог бы быть вполне достаточным не только для эволюции разума, но и для появления цивилизации. Заметим, что для развития человеческой цивилизации (начиная с появления примитивного социума человекообразных обезьян) потребовалось не более 4 – 6 млн. лет. Мы вовсе не утверждаем, что такое явление было в действительности, однако полностью исключить его нельзя.
2) «МИТОХОНДРИИ У ЧУЖИХ ОТСУТСТВУЮТ, И КОНЕЧНЫЕ СТАДИИ ОКИСЛИТЕЛЬНОГО МЕТАБОЛИЗМА ЭНЕРГЕТИЧЕСКИХ СУБСТРАТОВ ПРОИСХОДЯТ В ОСОБЫХ ИНТЕРДИГИТАЦИЯХ ПЛАЗМАЛЕММЫ - ТАК НАЗЫВАЕМЫХ СЕТЧАТЫХ ТЕЛЬЦАХ». Это очень странно, поскольку митохондрии присутствуют в клетках всех эукариотов, как в животных, так и в растительных. Сетчатые тельца представляют собой «...ДЛИННЫЕ ПАЛЬЦЕОБРАЗНЫЕ ВПЯЧИВАНИЯ ПЛАЗМАЛЕММЫ» и «... МОГУТ ТЯНУТЬСЯ ... К ОРГАНЕЛЛАМ, АКТИВНО РАСХОДУЮЩИМ ЭНЕРГИЮ...» . Из этих утверждений совершенно неясно, где именно локализованы медиаторы окислительного фосфорилирования. Известно, что обычная митохондрия окружена , двойной мембраной и что элементы электрон-транспортной цепи располагаются на ее внутренней мембране. Внешняя и внутренняя мембраны митохондрий имеют разное филогенетическое происхождение: считается, что появление митохондрий в клетках эукариотов связано с проникновением в них прокариотических клеток и дальнейшим развитием симбиоза. Внешняя мембрана митохондрий образовалась путем впячивания клеточной мембраны (плазмалеммы), а внутренняя была непосредственно мембраной симбионта. Даже в составе клеток современных эукариотов митохондрии сохранили известную самостоятельность, поскольку часть мембранносвязанных белков, играющих определенную роль в электронном транспорте кодируется непосредственно митохондриальной ДНК и находится на поверхности ее внутренней мембраны. В этой связи непонятно, каким образом впячивания самой плазмалеммы могут выполнять такую сложную функцию.
«ДОНОРОМ ЭЛЕКТРОНОВ СЛУЖИТ ВОССТАНОВЛЕННАЯ ФОРМА ОДНОГО ИЗ ХИНОИДНЫХ КОФЕРМЕНТОВ, А КОНЕЧНЫМ АКЦЕПТОРОМ - КИСЛОРОД. ПРИ ПЕРЕНОСЕ ОДНОЙ ПАРЫ ЭЛЕКТРОНОВ С ХИНОИДНОГО КОФЕРМЕНТА НА КИСЛОРОД ВО ВНЕКЛЕТОЧНОЕ ПРОСТРАНСТВО ВЫВОДИТСЯ ПЯТЬ ПАР ПРОТОНОВ. ПРОТОНЫ ВОЗВРАЩАЮТСЯ ВНУТРЬ КЛЕТКИ ЧЕРЕЗ ОСОБЫЙ ИОННЫЙ КАНАЛ (ПРОТОНЗАВИСИМУЮ ГТФ-СИНТЕТАЗУ)...». Здесь, конечно, можно провести параллель с дыхательной цепью митохондрий. Компоненты дыхательной цепи катализируют перенос электронов от восстановленного Никотинамидадениндинуклеотида (НАДН) или восстановленного убихинона на молекулярный кислород. Из-за большой разности окислительно-восстановительных потенциалов донора и акцептора реакция является высокоэкзергонической. Большая часть выделяющейся при этом энергии используется для создания градиента протонов и, наконец, для образования АТФ с помощью АТФ-синтазы.
«ГУАНОЗИНТРИФОСФАТ (ГТФ) - ОСНОВНОЕ МАКРОЭРГИЧЕСКОЕ СОЕДИНЕНИЕ». У эукариот, как растительных, так и животных, основную роль играет Аденозинтрифосфат (АТФ). В то же время, ГТФ важен в некоторых процессах, например, в цикле Кребса, при передаче сигнала от рецепторов мембраны к ядру клетки и др.
3). Кровь насекомых называется гемолимфой, т.к. их незамкнутая сосудистая система сообщается с межклеточным пространством. По-видимому, незамкнутая кровеносная система не функциональна для крупных животных, а малый рабочий Чужих, исходя из данных доклада имеет весьма внушительные размеры (более 2.5 м); по-видимому, боевые и высшие особи могут быть еще крупнее. Соответственно в тексте находим, что «КРОВЕНОСНАЯ СИСТЕМА ЧУЖИХ - ЗАМКНУТОГО ТИПА». Показательно, что кровь Чужих все-таки не названа гемолимфой. Если бы доклад являлся компиляцией какого-либо текста из области физиологии беспозвоночных, наверняка бы присутствовал бы термин «гемолимфа» вместо «кровь».
«КРОВЬ ЧУЖИХ - ПОЛУПРОЗРАЧНАЯ ОПАЛЕСЦИРУЮЩАЯ ЖИДКОСТЬ, ТЕМНО-СИНЯЯ В ОКСИГЕНИРОВАННОМ И СИНЕ-ЗЕЛЕНАЯ В ДЕЗОКСИГЕНИРОВАННОМ СОСТОЯНИИ... ГАЗОТРАНСПОРТНАЯ ФУНКЦИЯ ОПОСРЕДОВАНА КУПРОПРОТЕИДОМ АЛИЕНОЦИАНИНОМ, СОДЕРЖАЩИМ ЧЕТЫРЕ АТОМА МЕДИ НА ОДНУ МОЛЕКУЛУ. ОН ОБУСЛАВЛИВАЕТ ЦВЕТ КРОВИ ЧУЖИХ». Нужно сказать, что такой белок неизвестен. По-видимому, за неимением подходящего термина, это название было составлено из двух слов АЛИЕН (Чужой) и ЦИАНИН. Однако, совершенно очевидно, что речь здесь идет о ГЕМОЦИАНИНЕ. Гемоцианин – дыхательный пигмент из группы хемопротеидов, способный легко присоединять и отдавать кислород, является медьсодержащим аналогом гемоглобина, и встречактся в гемолимфе некоторых беспозвоночных. Молекула гемоцианина может содержать разное количество субъединиц. Обычно одна субъединица несет 2 атома меди, которые связывают одну молекулу кислорода. Таким образом, молекула гемоцианина, о котором идет речь в тексте, содержащая 4 атома меди, должна состоять из двух субъединиц.
Следует отметить, что гемоглобин (или его аналоги) - это очень важный таксономический признак. Его эволюция прослеживается на протяжени всего филогенетического развития организмов. Так фрагменты генов гемоцианина и его аналогов сейчас с успехом используются для уточнения таксономических субъединиц, последовательности филогенетического развития а также приблизительной оценки времени дивергенции родственных форм беспозвоночных [Burmester T. et al. THE EVOLUTION OF HEXAMERINS AND THE PHYLOGENY OF INSECTS. J Mol Evol, 1998, 47, :93–108; Kusche K. COMPLETE SUBUNIT SEQUENCES, STRUCTURE AND EVOLUTION OF THE 6 • 6-MER HEMOCYANIN FROM THE COMMON HOUSE CENTIPEDE, Scutigera coleoptrata. Eur. J. Biochem. 2003, 270, 2860–2868]. Наиболее древний гемоцианин имел 6-единичную структуру. В процессе эволюции членистоногих (хелицеровых, паукообразных) происходило формирование мультимеров (2 х 6, 4 х 6, 8 х 6 –меров - 540 - 420 миллионов лет назад). До последнего времени считалось, что гемоцианин ракообразных (Crustacea) и гексамерин насекомых (Insecta) имеет общее происхождение, но гексамерин утратил способность связывать медь и превратился в запасной белок (предполагалось, что насекомые могут обходиться без специального белка-транспортера кислорода. Доставка кислорода к тканям, якобы, происходит у них за счет большого количества разветвленных трахеол, кроме того, небольшое количество кислорода растворено непосредственно в гемолимфе; трудно представить, существовал ли такой механизм у гиганских вымерших форм). Время дивергенции гемоцианина и гексамерина представлялось гораздо более давним, чем расхождение самих классов [Burmester T. EVOLUTIONARY HISTORY AND DIVERSITY OF ARTHROPOD HEMOCYANINS.Micron, 2004, 35, 121–122]. Обычно гексамерин ракообразных представлен мономером 1 х 6 субъединиц или димером 2 х 6 субъединиц. Однако, недавно полученные данные показывают, что в организме некоторых насекомых все-таки может присутствовать гемоцианин. Причем обнаруженный гемоцианин имел 2-х субъединичную структуру (или 4 атома меди в молекуле). Этот факт был впервые подтвержден в статье, поданной в печать в 2003 г. [Hagner-Holler S. et al. A RESPIRATORY HEMOCYANIN FROM AN INSECT. PNAS, 2004, 101, 3, 871–874]. Материал был получен при исследовании гемолимфы одного из видов веснянки (Perla marginata). Следует отметить, некоторую таксономическую близость веснянок и термитов, т.к. они относятся к одной подкогорте Polyneoptera.
Таким образом, факт существования гемоцианина содержащего 4 атома меди в молекуле у насекомых до 2003 года не был известен даже узкому кругу специалистов. С другой стороны, как нам удалось выяснить, текст обсуждаемого доклада появился в интернете в 1996 году. Исходя из этого можно допустить, что обнаружение подобного «алиеноцианина» могло быть следствием непосредственного практического исследования, а отнюдь не литературных «изысканий».
Гемоцианин (в отличие от гемоглобина позвоночных, заключенного в эритроцитах) присутствует в гемолимфе членистоногих в растворенном состоянии или адсорбирован на поверхности клеток. У Чужих совершенно аналогичная ситуация: «АЛИЕНОЦИАНИН ФОРМИРУЕТ НАДМОЛЕКУЛЯРНЫЕ КОМПЛЕКСЫ С ЛИПИДАМИ ПЛАЗМЫ И СОРБИРУЕТСЯ НА ПОВЕРХНОСТИ ФОРМЕННЫХ ЭЛЕМЕНТОВ...».
4). «СУБМЕМБРАННЫЕ ТЕЛЬЦА ВСЕХ КЛЕТОК ЧУЖИХ СОДЕРЖАТ ТЕРМОСТАБИЛЬНЫЙ АКТИВАТОР КОАГУЛЯЦИИ, ВЫСВОБОЖДАЮЩИЙСЯ ПРИ ПОВРЕЖДЕНИИ ПЛАЗМАЛЕММЫ.». Термостабильный активатор коагуляции у насекомых неизвестен. Однако он «РАСЩЕПЛЯЕТСЯ ЛИПАЗОЙ ПЛАЗМЫ», т.е. должен быть липидной природы или липидным комплексом белка. Согласно предложенному механизму свертывания у инсект, отрицательно заряженные липиды, выделяемые гемоцитами играют определенную роль в начале процесса свертывания у дрозофилы [Theopold U. , Schmidt O. HELIX POMATIA LECTIN AND ANNEXIN V, TWO MOLECULAR PROBES FOR INSECT MICROPARTICLES:POSSIBLE INVOLVEMENT IN HEMOLYMPH COAGULATION. J. Insect Physiol. 1997,43, 667–674]. С другой стороны, название, возможно, отражает принадлежность фактора к так наз. heat-shock protein family [MacRae T. H. STRUCTURE AND FUNCTION OF SMALL HEAT SHOCK/ALPHA-CRYSTALLIN PROTEINS: ESTABLISHED CONCEPTS AND EMERGING IDEAS. Cell. Mol. Life Sci.2000, 57, 899–913].
«ТЕРМОСТАБИЛЬНЫЙ АКТИВАТОР КОАГУЛЯЦИИ...ФИКСИРУЕТСЯ В МЕСТАХ ПОВРЕЖДЕНИЯ СОСУДИСТОЙ СТЕНКИ И ЗАПУСКАЕТ ЦЕПЬ ФЕРМЕНТАТИВНЫХ РЕАКЦИЙ МЕЖДУ СПЕЦИФИЧЕСКИМИ БЕЛКАМИ ПЛАЗМЫ, ВЕДУЩУЮ К ОБРАЗОВАНИЮ ПЛОТНОГО СГУСТКА» Один из путей свертывания гемолимфы насекомых действительно предполагает существование каскада активации протеолитических ферментов, приводящего к образованию сгустка. Природа факторов и функционирование системы коагуляции и фибринолиза сейчас активно изучаются [Scherfer C. et al. THE TOLL IMMUNE-REGULATED DROSOPHILA PROTEIN FONDUE IS INVOLVED IN HEMOLYMPH CLOTTING AND PUPARIUM FORMATION. Dev Biol., 2006, 295, 1,156-163]. Таким образом в описании системы коагуляции «Чужих» и насекомых есть некоторые соответствия, но система коагуляци насекомы, изучается давно (с 1954 г), поэтому трудно понять, является ли описание этой системы в тексте цитированием.
5) Далее хотелось бы остановиться немного на отдельных моментах.
«МОЧЕВАЯ КИСЛОТА - КОНЕЧНЫЙ ПРОДУКТ АЗОТИСТОГО ОБМЕНА». У птиц, ряда пресмыкающихся и большинства наземных насекомых мочевая кислота - конечный продукт не только пуринового, но и белкового обмена. Система биосинтеза мочевой кислоты (а не мочевины, как у большинства позвоночных) служит для связывания аммиака, токсичного продукта азотистого обмена, и развилась у этих животных в связи с характерным для них ограниченным водным балансом (выводится из организма с минимальным количеством воды или даже в твёрдом виде).
«МИОЭПИТЕЛИЙ ТОНКОЙ КИШКИ СЕКРЕТИРУЕТ СМЕСЬ ГИДРОЛАЗ С PH-ОПТИМУМОМ В НЕЙТРАЛЬНОЙ ОБЛАСТИ» - известны несколько протеаз с нейтральным рН оптимумом – трипсин, например. Пепсин и эластаза, повидимому, отсутствуют, т.к. их рН оптимумы составляют, соответственно, 1-2 и 8-9..
В целом доклад написан очень грамотно и, возможно, является результатом прямых исследований. Однако, настораживает огромный объем представленного материала. Маловероятно, чтобы небольшая группа ученых могла выполнить исследования, которые по плечу только ведущим научно-исследовательским институтам, причем с объемом работы для десятка институтов не менее, чем на 2 – 3 десятилетия. Впрочем, в бывшем СССР существовали потенциальные возможности для таких исследований. И за интересующий период (с 1965 по 1996 гг.) времени было достаточно.
Автор: Э.Н. Золотарева, кандидат биологических наук.
-------------------------------
Мнение доцента Б.Ф.Иванова, сотрудника кафедры биологии Воронежского Государственного Университета:
"Пpочтение данного файла заняло довольно много времени т.к. не хотелось читать "по диагонали". Общее впечатление: безусловно грамотно описанный обьект. Hо вот какой? Если отбросить упоминания о "чужих" и некоторую экзотику, то анатомический раздел похож на выписку из научного отчета или диссертации, посвященной какому-то довольно редкому, но подозрительно "земному" созданию. Hапрашивается мысль, что "чужие" о которых идет речь не инопланетяне, а нечто найденное в каком-нибудь изоляте вполне на Земле. Слишком легко ложится все в общепринятую терминологию. IMHO, собственно инопланетные существа должны были бы поставить анатомов в большой тупик своей несхожестью с известными формами. Так что, в данном случае, профессионализм описания скорее настораживает, чем убеждает. Hапрашивается мысль о розыгрыше с использованием готового и вполне серьезного материала, но написанного по совсем другому поводу. М. б. компилляция. Это вовсе не потребовало бы того количества времени и пива, которое предполагают скептики.
С другой стороны, зоологи, читавшие данный текст, не могут указать подходящий обьект к которому все написанное может быть отнесено. Вполне возможно, что тексту уже много лет и сам он является продуктом довольно долгой работы, так что когда его составляли, терминологические трудности уже были преодолены. Или авторы сознательно не акцентировали непонятных для них вещей и все уложили в привычные слова и формулировки. Hо в любом случае употребление терминов из привычной "земной" систематики и гистологии настораживает. Хотя это только мое личное мнение.
Hаиболее понятную для меня часть текста - физиолого-биохимическую писал явно не профан. Hо: изложенная в этом разделе информация потребовала бы такого огромного обьема работы и привлечения специалистов такого класса, что не верится в возможность удержать все это в секрете. Одна только проблема биоэнергетики - сетчатые структуры вместо митохондрий - тянет на Hобелевскую премию. А там еще и ионные каналы и нетипичные коферменты и алиеноциан... Это работа на много лет нескольким институтам. И армии вертухаев вокруг. С другой стороны, тут некоторые вещи "недостаточно безумны". Hу, скажем, подразумевающееся совпадение аминокислотного состава чужих с земными организмами... В синтез щавелевоуксусной кислоты из двууглеродных соединений верится, а вот с хромосомами - какая-то невнятица. И вообще, весь раздел как то напоминает переписанный озорником учебник. Грамотным озорником, впрочем.
Среди вещей, похожих на проколы отмечу:
> магнитное поле мембраны _и_ > ЦЕРЕБРАЛЬHЫЙ МАГHИТОСЕHСОРHЫЙ АППАРАТ *который* ЯВЛЯЕТСЯ ОСHОВHЫМ > ОРГАHОМ ДИСТАHТHОЙ РЕЦЕПЦИИ МАЛОГО РАБОЧЕГО. *причем* ПРИHЦИП РАБОТЫ > ДАHHОГО ОРГАHА ЧУВСТВ - РЕГИСТРАЦИЯ ПЕРЕМЕHHОГО МАГHИТHОГО ПОЛЯ > СЛОЖHОЙ КОHФИГУРАЦИИ, ГЕHЕРИРУЕМОГО САМИМ АППАРАТОМ
Вообще же, о работе ЦHС чужих известно подозрительно много. О ЦHС человека и то так уверенно не заявляют касательно механизмов хранения памяти. Разве что сами чужие просветили на этот счет... Что не является таким уж диким предположением.
Полагать данный текст подлинным означает признать, что относительно недавно - не раньше семидесятых годов - где-то проводилась огромная научная работа с десятками особей чужих. Я могу поверить в посещение Земли инопланетянами или даже наличие на ней скрытой от нас формы разумной жизни, но в способность любого государства удержать в секрете исследования такого масштаба верится с большим трудом. Хотя, вспоминая разные лабораторные сплетни, не могу полностью отрицать этот вариант.
Ну,уважаемая Нефелана,Вы мне здорово польстили)).Читала этот материал давно,частично еще в сканированном виде .Думаю,написано в конце 60-70е годы.Общее впечатление: анализируется реальный объект неземного происхождения,хотя я не биолог,а врач.Точно одно-писал профессиональный анатом с хорошим биологическим образованием. Профессионалы такого уровня крайне редко увлекались фантастикой,М.Попович редчайшее исключение,в основном это было люди суровые,мало склонные к мистификациям))).Учитывая,что материал появился еще до развития общемировых эсхатологических ожиданий, это может быть правдой.Обратите внимание на отсутствие митохондрий,у Радмара есть отдельная тема о митохондриях. Утечка возможна всегда и отовсюду,немцы говорят:" что знают двое,знает свинья " .Не знаю,как Вы,а я,грешная,если бы попала в такую лабораторию,обязательно бы скопировала документы на всякий случай для себя А потом всякое могло быть,вдруг автор умер,и наследники откопали случайно,или еще какой вариант... В целом:я -за подлинность.
Сообщение отредактировал джекки - Пятница, 14.01.2011, 18:03
ГИСТОЛОГИЧЕСКОЕ И ЭЛЕКТРОННО-МИКРОСКОПИЧЕСКОЕ ИССЛЕДОВАНИЕ ПЕЧЕНИ ЯЩЕРИЦ С БЫВШЕГО СЕМИПАЛАТИНСКОГО ИСПЫТАТЕЛЬНОГО ПОЛИГОНА (ВОСТОЧНЫЙ КАЗАХСТАН)
Т.Н. Дуйсебаева 1, Г.В. Федотовских 2, Н.А. Стрелюхина 3 1 Институт зоологии МОН РК Казахстан, 050060, Алматы, пр. Аль-Фараби, 93 2 Научный Центр хирургии им. Сызганова Казахстан, 050004, Алматы, пр. Желтоксан, 62 3 Институт экспериментальной биологии МОН РК Казахстан, 050030, Алматы, пр. Абая, 78 Поступила в редакцию 16.02.2007 г.
ВВЕДЕНИЕ
В литературе мало сведений по воздействию радиационного излучения на организм амфибий, рептилий и птиц. В основе большинства работ по изучению отдаленных последствий ионизирующей радиации на живой организм лежат экспериментальные исследования на млекопитающих (Кропачева, Мишурова, 1988; Богатов и др., 1991; Костеша, Лопухова, 1991). После чернобыльской катастрофы 1985 г. амфибии и рептилии также явились объектами серии исследований по воздействию на живой организм малых доз радиации низких интенсивностей. Однако работы с этими позвоночными ограничились в основном изучением содержания радионуклидов во внутренних органах, цитогенетическим мониторингом и анализом структуры популяций (Елисеева и др., 1994, 1995; Пикулик, Дробенков, 1995; Пикулин, Пленин, 1995). Те же задачи преследовало изучение бесхвостых амфибий на территории Восточно-Уральского радиационного следа (Черданцев и др., 1993). Отдельные цитологические нарушения, выражающиеся в деструкции гепатоцитов, гипертрофии клеток гемопоэтического окружения, увеличении размеров ядер и снижении митотической активности, были отмечены только для печени лягушек (Пястолова и др., 1996; Пястолова, Вершинин, 1999). К сожалению, в доступной нам литературе мы не встретили работ по цитологии и ультраструктуре печени у пресмыкающихся, населяющих территории, пострадавшие от радиационного воздействия.
Бывший Семипалатинский испытательный полигон (СИП), расположенный в Восточном Казахстане, относится к одной из уникальных точек на планете, являясь природной радиоэкологической лабораторией. Многолетнее воздействие ионизирующей радиации не могло не отразиться на сообществах обитающих здесь растений и животных. В настоящее время комплексные исследования на биологических объектах полигона проводятся в ряде научных учреждений Казахстана. Из зоологических объектов традиционными продолжают оставаться млекопитающие, как наиболее радиочувствительные животные (Черных и др., 1998, 1999), хотя морфологические и цитогенетические исследования начаты и для других групп позвоночных (Атаханова, 1998; Ильинских и др., 1998; Крайнюк, 1998). Располагая материалом по двум видам ящериц с территории СИП и опираясь на результаты радиационных экспериментальных работ по печени амфибий и млекопитающих, мы поставили целью настоящего исследования изучение эффектов радиационного воздействия разной дозы на внутреннюю и тонкую структуру печени ящериц, постоянно обитающих на территории бывшего СИП.
МАТЕРИАЛ И МЕТОДЫ
Материалом для настоящего исследования послужила печень 9 взрослых особей прыткой ящерицы (Lacerta agilis) с длиной тела 54 – 90 мм и 14 взрослых особей разноцветной ящурки (Eremias arguta) с длиной тела 52 – 79 мм. Животные были отловлены в летний период 1999 – 2001 гг. на территории бывшего СИЯП, а также в других районах Восточного Казахстана с уровнем радиации в пределах нормы. Ниже приведены точки сбора материала с указанием мощности экспозиционной дозы (МЭД, мкР/ч). Данные МЭД получены с использованием показаний индивидуального дозиметра типа ДЛТ-03 при непосредственном сборе материала и уточнены по литературным данным (Апсаликов и др., 1996): 1) подножье гор Семейтау, МЭД 10 – 12 мкР/ч (фоновый уровень), L. agilis (n = 4), E. arguta (n = 3); 2) западный берег р. Чаган, 1 – 1.5 км южнее оз. Чаган, СИП, МЭД 100 – 200 мкР/ч, L. agilis (n = 3), E. arguta (n = 5); 3) западный берег оз. Чаган (края воронки), СИП, МЭД 400 – 1500 мкР/ч, L. agilis (n = 2), E. arguta (n = 6).
По ряду причин определение суммарной дозы облучения (внешнего и внутреннего) ящериц затруднительно (см. Апсаликов и др., 1996: 127 – 128). Поэтому в работе мы приводим только значения мощности экспозиционных доз для соответствующих местностей без расчета суммарной дозы облучения, полученной животными. Очевидно, что приуроченные к обитанию на ограниченной территории (индивидуальный участок площадью около 40×40 м2) и не способные к протяженным миграциям особи из мест обитания с разной МЭД будут соответственно различаться по величине полученной дозы облучения.
Для гистологического изучения кусочки печени исследуемых животных, собранных в точках 2 (МЭД 100 – 200 мкР/ч) и 3 (МЭД 400 – 1500 мкР/ч), фиксировали в 10% нейтральном формалине. После обработки по общепринятой методике и заливки в парафин-воск срезы (5 – 7 мкм) окрашивали гематоксилином и эозином (Волкова, Елецкий, 1971). Полихромная окраска по Массону (Martoja, MartojaPierson, 1967) использовалась, главным образом, для выявления соединительной ткани. Для электронно-микроскопического исследования биопсийные кусочки ткани животных, собранных в точке 3 (МЭД 400 – 1500 мкР/ч), фиксировали в 2.5% растворе глютаральдегида на 0.2M фосфатном буфере Миллонига (рH 7.4 – 7.6) в течение 2 часов с постфиксацией в 1% растворе четырехокиси осмия (2 часа). Проводка осуществлялась по общепринятой методике с заключением в эпоксидную смолу эпон. Полутонкие и ультратонкие срезы готовились на ультрамикротомах Reichert OM U2 и Nova LKB-3. Полутонкие срезы, толщиной около 3 микрон, окрашивали полихромным красителем – метиленовым синим, азуром-2 и основным фуксином по Humphrey и Pittman (1974). Ультратонкие срезы толщиной около 600 ангстрем контрастировали 2.5% раствором уранилацетата и цитратом свинца по Reynolds (1963). Ультратонкие срезы изучали в электронном микроскопе ЭВМ-100.
РЕЗУЛЬТАТЫ
Гистологическое исследование печени ящериц Lacerta agilis и Eremias arguta
Гистологическое исследование показало, что у изученных ящериц из контрольной группы гепатоциты имели полигональную форму с округлым ядром в центральной части. Соединительная ткань в междольковых перегородках не выявлялась, и границы печеночных долек определялись только по скоплениям сосудов, располагающихся на границе соседних печеночных долек. В цитоплазме гепатоцитов встречались мелано-макрофагальные комплексы. Небольшое их количество обнаруживалось и в глиссоновой капсуле.
У ящериц обоих видов из опытной группы морфологические изменения в печени были сходными, поэтому мы приводим результаты исследования независимо от видовой принадлежности особей. У ящериц, обитающих в зоне МЭД 100 – 200 мкР/ч, обращало на себя внимание неравномерное полнокровие печени. Большинство капилляров, венул и синусоидов были расширены и полнокровны, в то время как другие малокровны. Иногда можно было видеть старые и свежие очаговые и диапедезные кровоизлияния, лимфоцитарные выпоты, явления периваскулярного отека (рис. 1).
Рис. 1. Гистология печени прыткой ящерицы в опытной группе. Полнокровие синусоида и капилляра, вакуольная дистрофия гепатоцитов. МЭД 100 – 200 мкР/ч. Окраска гематоксилином и эозином. Ув. × 250
В очагах старых кровоизлияний ядра эритроцитов были лизированы. Дистрофические изменения в печени носили характер вакуольной и жировой дистрофии. Явления вакуольной дистрофии были выражены в центральных частях долек, однако не во всех из них одинаково интенсивно. Жировая дистрофия гепатоцитов локализовалась по периферии дольки (рис. 2).
Иногда встречались очаги некробиоза и некроза. В очагах микронекроза ядра подвергались лизису, реже пикнозу и рексису. Во многих клетках ядра не были видны, и вся клетка имела вид тусклого глыбчатого образования. Некоторые очажки микронекроза имели вид небольших зернистых, сравнительно четко контурированных участков, в которых отдельные гепатоциты не различались. По периферии и внутри очажков можно было видеть инфильтрацию лимфоцитов, гистиоцитов и купферовских клеток. Местами встречалась очаговая дискомплексация гепатоцитов. Количество двуядерных гепатоцитов увеличивалось, по сравнению с контролем (МЭД 10 – 12 мкР/ч). Нередко в ядрах наблюдалось по два гипертрофированных ядрышка.
В печени ящериц, собранных при мощности экспозиционной дозы 400 – 1500 мкР/ч, нарушения кровообращения нарастали: капилляры, венулы и синусоиды были расширены и переполнены кровью, увеличивались отеки и количество очагов микронекрозов. Купферовские клетки нередко были гиперплазированы, некоторые из них подвергались некрозу. Отмечалось также очаговое слущивание эндотелиоцитов в просветы синусоидов. Внутри долек местами встречались мелкие скопления лимфоцитов. В периваскулярных пространствах иногда можно было видеть отечную жидкость, а в стенках отдельных сосудов были отмечены явления микрофокусного плазматического пропитывания. Стенки отдельных сосудов были утолщены и разволокнены. Часто встречались свежие и старые очаговые, периваскулярные и диапедезные кровоизлияния.
Изменения гепатоцитов были представлены вакуольной и жировой дистрофией. Очаги микронекрозов и дискомплексация гепатоцитов отмечались чаще, чем у ящериц, обитающих в зоне при 100 – 200 мкР/ч. Гепатоциты в состоянии вакуольной дистрофии были набухшие, ядро их было нередко смещено к периферии (эктопия ядер). Ядра большинства гепатоцитов были крупные, просветленные, но встречались темные ядра, иногда пикнотичные. Гепатоциты с темными уменьшенными ядрами встречались преимущественно в периваскулярных зонах ацинусов.
Также обращало на себя внимание значительное увеличение числа двуядерных гепатоцитов. Встречалось множество гепатоцитов с крупными ядрами, содержащими по 2 или 4 ядрышка. Выявлялись и гепатоциты с базофильной цитоплазмой.
Электронно-микроскопическое исследование печени ящериц Lacerta agilis и Eremias arguta
Ультраструктура печени прыткой ящерицы в контрольной группе. Морфологическое исследование полутонких срезов печени на светооптическом уровне выявило неоднородную структуру ткани, представленную традиционными трубчатыми структурами из крупных гепатоцитов на фоне широких синусоидов и полями мелких, разделенных тонкой ретикулярной стромой и значительно удаленных от узких синусоидов, гепатоцитов.
На электронно-микроскопическом уровне характерной особенностью гепатоцитов была ярко выраженная депонирующая функция. Большая часть гиалоплазмы была заполнена электронно-плотными розетками и гранулами гликогена, а также полиморфными по величине осмиофильными липидными включениями (рис. 3).
Крупные и плотные митохондрии с обвивающими их канальцами гранулярного эндоплазматического ретикулума располагались возле ярда и по периферии гепатоцитов. В области желчных капилляров, сформированной обычно четырьмя гепатоцитами, располагались элементы комплекса Гольджи, пузырьки и крупные эндоплазматические ретикулумы и многочисленные цитосегресомы, представленные миелиноподобными фигурами с осмиофильными зернистыми вкраплениями типа ферритина. Синусоидальная поверхность гепатоцитов содержала микроворсинки и характеризовалась на отдельных участках выраженным микропиноцитозом. Базальная мембрана имела прерывистый вид. Пространство Диссе было заполнено хлопьевидным материалом и нежными волоконцами тропоколлагена со слабовыраженной поперечной исчерченностью. Такие приспособительные реакции, как клазматоз цитоплазмы и активное высвобождение гепатоцитов от цитосегресом через синусоидальную поверхность, отражали высокий уровень обменных процессов в печени. Цитосегресомы фагоцитировались эндотелиальными клетками синусоидальной выстилки.
Наряду с клетками Купфера нами были отмечены немногочисленные малодифференцированные пигментные клетки с крупными премеланосомами.
Рис. 3. Ультраструктура печени прыткой ящерицы в контрольной группе. Цитосегресомы и крупные липидные капли в цитоплазме крупного гепатоцита. Ув. ×12000
Ультраструктура мелких гепатоцитов отличалась слабовыраженной депонирующей функцией. Гиалоплазма данных клеток была равномерно заполнена немногочисленными гранулами гликогена. Липидные включения были скудными. Небольшие и плотные митохондрии, каналы гранулярного эндоплазматического ретикулума располагались без определенной закономерности. Помимо цитосегресом и крупных фаголизисом, в цитоплазме мелких гепатоцитов были отмечены свободнолежащие в гиалоплазме электронно-плотные зерна гемосидерина. Плотно прилежащие друг к другу гепатоциты соединялись межклеточными контактами типа замка. Пласты гепатоцитов были окружены утолщенной базальной мемебраной. Широкие межклеточные щели были заполнены элементами ретикулярной стромы и образующимися в процессе клазматоза фрагментами гепатоцитов. В просвете синусоидов были часто видны распадающиеся клетки Купфера.
Таким образом, печень L. agilis в контрольной группе исследования характеризовалась «рекомбинацией» гепатоцитов, являющейся морфологическим эквивалентом приспособительных реакций, приобретенных в филогенезе и связанных с определенными условиями функционирования (Калашникова, 1996).
Ультраструктура печени прыткой ящерицы в опытной группе. В опытной группе ящериц L. agilis на фоне преобладающего большинства гепатоцитов с высоким, как и в контроле, уровнем депонирующей функции, нами были отмечены гепатоциты с ярко выраженными признаками гликогенолиза и уменьшения липидных включений. Резкое снижение электронной плотности гранул гликогена проявлялось на светооптическом уровне как бы лакированной розовой окраской цитоплазмы гепатоцитов. Сохранялась лишь небольшая зона возле ядра, занятая немногочисленными органеллами и рибосомами.
Наблюдалось также некоторое увеличение числа цитосегресом и зернообразных включений как вкрапленных в миелиноподобные фигуры цитосегресом, так и свободно расположенных в гиалоплазме (рис. 4).
Рис. 4. Ультраструктура печени прыткой ящерицы в опытной группе. Многочисленные цитосегресомы с зернами типа ферритина. Ув. ×12000
На отдельных, более бледных на светооптическом уровне участках печени, к данным изменениям присоединялись явления вакуолизации агранулярного эндоплазматического ретикулума, образование крупных баллонов, заполненных отечной жидкостью. Деструкция канальцев ретикулума приводила к очаговому оводнению гиалоплазмы и появлению многочисленных тонких миелиноподобных фигур, образующихся в результате вторичной структуризации фосфолипидов. В состоянии вакуолизации и дегрануляции мембраносвязанных рибосом находились и канальцы гранулярного эндоплазматического ретикулума. В отдельных гепатоцитах располагались значительные скопления крупных вторичных лизосом, формирующихся на месте деструктивно измененных электронно-плотных митохондрий и фрагментов гранулярного эндоплазматического ретикулума.
В цитоплазме клеток Купфера располагались крупные обломки фагоцитированных эритроцитов, формирующие массы гемосидерина с миелиноподобными структурами по периферии, а также мелкие электронно-плотные гранулы. В цитоплазме пигментных клеток были отмечены многочисленные везикулы комплекса Гольджи, трубочки агранулярного эндоплазматического ретикулума, премеланосомы и зрелые электронно-плотные гранулы меланина.
Ультраструктура мелких гепатоцитов характеризовалась накоплением электронноплотных масс гемосидерина и усиленным клазматозом цитоплазмы, высоким уровнем ядерно-белкового и энергетического синтеза. Ультраструктура печени разноцветной ящурки в опытной группе. Электронно-микроскопическое исследование печени разноцветной ящурки вне очагов микронекроза показало значительное накопление крупных электронно-плотных масс гемосидерина в цитоплазме гепатоцитов (рис. 5).
Рис. 5. Ультраструктура печени разноцветной ящурки в опытной группе. Крупные массы гемосидерина в гиалоплазме гепатоцитов. Ув. ×12000
Отмечена трансформация липидных капель в гранулы гликогена. На месте резорбирующихся липидных включе- ний оставались миелиноподобные мембранные структуры и небольшие очаги просветления гиалоплазмы. Выраженная гиперплазия агранулярного эндоплазматического ретикулума была направлена на компенсаторный синтез липидных включений, мелкие новообразованные капли которых располагались возле мелких везикул агранулярного эндоплазматического ретикулума.
Характерной особенностью печени данных животных было наличие большого числа макрофагов моноцитного типа, незрелых и зрелых клеток Купфера, полностью заполняющих просвет синусоидов, и находящихся на различных этапах катаболизма погибающих фагоцитированных эритроцитов. Клетки Купфера содержали различные по размеру и структуре фагосомы. Наряду с обломками эритроцитов, электронноплотными массами с зернами ферритина, миелиноподобными структурами, связанными с перевариванием липидов стромы эритроцитов, в цитоплазме клеток Купфера появлялись и электронно-плотные гранулы меланина, непосредственно включенные в фагосомы (рис. 6).
Рис. 6. Ультраструктура печени разноцветной ящурки в опытной группе. Клетка Купфера с глыбками гемосидерина и миелиноподобными структурами. Ув. × 9800
Пигментные клетки содержали премеланосомы и гранулы меланина, свободно расположенные в гиалоплазме. Многие макрофаги находились в состоянии деструкции. Те же виды фагосом, отмеченные нами в цитоплазме клеток Купфера, наблюдались и в гиалоплазме гепатоцитов.
Полученные нами ультраструктурные картины полностью подтвердили данные М.М. Калашниковой (2000) о том, что выходящие из распадающихся клеток Купфера фагосомы поступают в цитоплазму гепатоцитов и выделяются в желчь через желчные капилляры. В области комплекса Гольджи, расположенного в перибилиарной зоне, данные структуры, по-видимому, подвергаются ферментативной обработке и лизису, превращаясь в электронно-прозрачные пузыри, хорошо видимые на полутонких срезах. Высокий уровень обменных процессов обеспечивался усиленным ядерно-белковым синтезом и крупными электронно-плотными энергезированными митохондриями.
В отдельных гепатоцитах при еще большем напряжении катаболизма фагоцитированных эритроцитов, когда вдоль синусоидальной поверхности появлялись целые ряды мелких и крупных фагосом, а в перибилиарной зоне «дорожка» трансформируемых фагосом, нами было отмечено набухание митохондрий, свидетельствующих об активации фосфорилирующей функции. Высокий уровень белкового синтеза отражался в появлении полей гиперплазированных и расширенных канальцев гранулярного эндоплазматического ретикулума, заполненных хлопьевидным материалом. Перенапряжение энергообразующей функции митохондрий приводило к фрагментации и лизису крист митохондрий. В участках, прилежащих к очагам микронекрозов, гепатоциты находились в состоянии парциального некроза.
ОБСУЖДЕНИЕ
Печень является великолепным индикатором состояния организма, поскольку весьма чувствительна к изменению внутренней среды организма или различного рода внешним факторам. Показано, что действие токсических веществ, в том числе при индустриальном загрязнении мест обитания, ведет к увеличению индекса печени, снижению цитоядерного соотношения гепатоцитов, увеличению размеров их ядер и появлению гигантских ядер (Пястолова, Трубецкая, 1989).
Результаты экспериментальных радиационных работ, проведенных на млекопитающих, показали, что после перенесения животными острой лучевой болезни в печени происходят серьезные деструктивные изменения, включающие в том числе нарушения гемодинамики, отложения в больших количествах гемосидерина, жировую и гидропическую вакуольную дистрофию, значительные изменения в ядрах и ядрышках гепатоцитов (Костеша, Лопухова, 1991).
В печени млекопитающих, подверженных экспериментальному облучению в малых дозах и с низкой интенсивностью, были выявлены значительные неспецифические изменения гепатоцитов, липидная, паренхиматозная и вакуольная дистрофии, некроз и гиперплазия купферовских клеток, развитие предопухолевых изменений в виде микрохолангиом и другие патологии (Пинчук и др., 1991).
Проведенное нами гистологическое исследование показало, что длительное воздействие ионизирующей радиации вызывало в печени ящериц заметные патоморфологические изменения, причем они были сходными у ящериц обоих видов (L. agilis, E. arguta) и зависели от мощности дозы радиации в зоне обитания животных.
Характерными изменениями были расстройства кровообращения в виде неравномерного кровенаполнения органа при МЭД 100 – 200 мкР/ч и венозного, синусоидального и капиллярного полнокровия, приводящих к гипоксии, отекам и кровоизлияниям при МЭД 400 – 1500 мкР/ч. Патологические изменения характеризовались также вакуольной и жировой дистрофией, очаговой дискомплексацией гепатоцитов, появлением очагов микронекрозов, в которых нередко отмечалась инфильтрация лимфоцитов, гистиоцитов и купферовских клеток, которая расценивается как реакция печени на повреждение.
Особенностью изменений в печени у ящериц из зоны МЭД 400 – 1500 мкР/ч являлось то, что наряду с выраженными гемодинамическими, дистрофическими и некротическими изменениями, наблюдались и компенсаторно-приспособительные процессы. Эти процессы выражались в появлении увеличенных гепатоцитов с базофильной цитоплазмой, значительном увеличении количества двуядерных гепатоцитов и появлением гепатоцитов с крупными ядрами, содержащими по 2 – 4 ядрышка. Увеличение ядер гепатоцитов при равных с контролем индексах печени, а также некоторые другие признаки деструкции гепатоцитов отмечались рядом авторов и в печени остромордых лягушек (Rana arvalis) из зоны радиационного загрязнения Чернобыля и Восточно-Уральского следа (Носова и др., 1994; Пястолова, Вершинин, 1999). Приведенные изменения свидетельствовали об усилении энергетических, гликогенообразовательных и белоксинтетических процессов в гепатоцитах в ответ на сильный экзогенный, а возможно, и эндогенный раздражитель (Саркисов, 1970).
Картина патологических изменений в печени ящериц при дозе 400 – 1500 мкР/ч, описанная в настоящей работе, оказалась весьма сходной с результатами гистологического исследования печени овец (Жапбасов и др., 1998), круглогодично содержавшихся в районе атомного озера «Балапан» на территории Семипалатинского полигона с заметно меньшим радиационным фоном (МЭД 25 – 100 мкР/ч).
В печени подопытных овец были отмечены расширения и отеки перикапиллярных пространств и синусоидов, встречалось большое количество гепатоцитов с признаками жировой и вакуольной дистрофии, скопления темных клеток, двуядерные гепатоциты и гигантские гепатоциты с крупными ядрами и множественными ядрышками. Электронно-микроскопическое исследование печени ящериц в опытной группе обнаружило появление признаков гликогенолиза и уменьшения липидных включений, явления гидропической дистрофии и внутриклеточного отека в определенной части гепатоцитов. Вне очагов микронекроза было отмечено значительное накопление гемосидерина в гепатоцитах в результате усиленного катаболизма погибших фагоцитированных эритроцитов. Высокий уровень обменных процессов обеспечивался утилизацией более эффективных в энергетическом отношении жировых включений, высоким уровнем ядерно-белкового синтеза и интенсификацией фосфорилирующей функции митохондрий. Функциональная активность гиперплазированных клеток Купфера была направлена на синтез пигментных гранул, важная роль среди которых отводится меланину как мощному антиоксиданту, защищающему организм от свободных радикалов кислорода.
ЗАКЛЮЧЕНИЕ
Все вышеизложенное позволяет сделать вывод о том, что длительное воздействие ионизирующей радиации в зоне обитания ящериц L. agilis и E. arguta приводит как к патологическим изменениям в печени этих пресмыкающихся (при мощности экспозиционной дозы 100 – 200 мкР/ч), так и к появлению компенсационно-приспособительных реакций, направленных на защиту организма от действия поражающего фактора (при мощности экспозиционной дозы 400 – 1500 мкР/ч).
Благодарности
Авторы благодарят Ж.Б. Левину (Казахстанский государственный университет им. Аль-Фараби, г. Алматы) и А.И. Жарикова (Научный центр хирургии им. Сызганова, г. Алматы) за техническую помощь в подготовке биопсийного материала для последующего гистологического и электронно-микроскопического изучения; И.Н. Магду (Институт зоологии, г. Алматы) за ценные консультации по радиационной обстановке на территории бывшего Семипалатинского полигона и В.А. Хромова (Семипалатинский государственный университет, г. Семипалатинск) за помощь при проведении полевых работ на территории бывшего СИЯП и ряд публикаций по радиационным исследованиям, предоставленным авторам для ознакомления.
Работа выполнена при финансовой поддержке фондов ИНТАС (проект INTAS-Kazakhstan 95-0016 «Environmental effects on amphibians and reptiles in severely polluted or destroyed habitats of Kazakhstan», 1997 – 1999) и НАТО (проект «Genetic screening of reptiles from Semipalatinsk «Polygon» nuclear test site», 2000 – 2001).
СПИСОК ЛИТЕРАТУРЫ Апсаликов К.Н., Гусев Б.И., Дусь В.И., Леонгардт Р.Б. Семипалатинское атомное озе- ро. Алматы: Гылым, 1996. 301 с. Атаханова К. Я. Эколого-генетическая характеристика некоторых видов позвоночных животных, обитающих на территории Семипалатинского ядерного полигона // Радиацион- ная безопасность и социально-экологические проблемы Казахстана. Караганда; Алматы: Гылым, 1998. С. 121 – 129. Волкова О.В., Елецкий Ю.К. Основы гистологии с гистологической техникой. М.: Ме- дицина, 1971. 272 с. Богатов Л.В., Сурина А.Г., Антоненко Г. И., Першина Л.И. Функциональное состояние щитовидной железы у овец в отдаленный период после однократного введения Sr90, Pu239 и их смеси // Радиобиология. 1991. Т. 31, вып. 4. С. 481 – 484. Елисеева К.Г., Войтович А.М., Плоская М.В., Смаль С.Э. Генетический мониторинг популяций бурых лягушек, обитающих в загрязненных радионуклидами районах Республи- ки Беларусь // Радиац. биол. Радиоэкология. 1994. Т. 34, вып. 6. С. 838 – 846. Елисеева К.Г., Войтович А. М., Трусова В.Д., Огурцова С.Э. Цитогенетический мони- торинг популяции амфибий // Животный мир в зоне аварии Чернобыльской АЭС. Минск: Навука i тэхнiка, 1995. С. 148 – 156. Жапбасов Р.Ж., Сейсебаев А.Т., Каримбаева А.Т., Карабалин Б.К., Тусупбаева В.И., Шеналь К., Нургалиева К.Ж. Цитогенетическое изучение млекопитающих, обитающих на территории бывшего Семипалатинского испытательного полигона // Радиационная безо- пасность и социально-экологические проблемы Казахстана. Караганда; Алматы: Гылым, 1998. С. 64 – 68. Ильинских Н.Н., Ксенз А.С., Ильинских Е.Н., Юркин А.Ю. Генотоксическая оценка со- стояния окружающей среды в зоне влияния Семипалатинского ядерного полигона // Радиа- ционная безопасность и социально-экологические проблемы Казахстана. Караганда; Алма- ты: Гылым, 1998. С. 117 – 121. Калашникова М.М. Особенности ультраструктуры клеток печени в сравнительно- морфологическом ряду животных и их значение // Бюл. экспериментальной биологии и ме- дицины. 1996. № 6. С. 604 – 609. Калашникова М.М. Ультраструктура клеток печени рыб и амфибий при катаболизме погибающих эритроцитов // Бюл. экспериментальной биологии и медицины. 2000. №1. С. 117 – 120. Костеша Н.Я., Лопухова В.В. Отдаленные эффекты у леченых собак, перенесших ост- рую лучевую болезнь // Радиобиология. 1991. Т. 21, вып. 4. С. 491 – 495. Крайнюк В.Н. Морфоэкологическая характеристика популяций карпа Cyprinus carpio L. водоемов территории Семипалатинского ядерного полигона // Радиационная безопас- ность и социально-экологические проблемы Казахстана. Караганда; Алматы: Гылым, 1998. С. 129 – 133. Кропачева К., Мишурова Е. Продолжительность скрытого повреждения печени после окончания хронического γ-облучения // Радиобиология. 1988. Т. 28, вып. 1. С. 44 – 47. Носова Л.И., Шалимов В.А., Рясенко В.И., Шевченко Ж.Т. Динамика изменения гема- тологических показателей у позвоночных зоны ЧАЭС // Итоги 8-летних работ по ликвида- ции последствий аварии на ЧАЭС. Зеленый Мыс, 1994. С. 253. Пикулик М.М., Дробенков С.М. Амфибии и рептилии // Животный мир в зоне аварии Чернобыльской АЭС. Минск: Навука i тэхнiка, 1995. С. 55 – 59. Пикулик М.М., Пленин А.Е. Предисловие // Животный мир в зоне аварии Чернобыль- ской АЭС. Минск: Навука i тэхнiка, 1995. С. 3 – 8. Пинчук В.Г., Никитченко В.В., Гольдшмидт В.Я., Андрущак Л.И., Серкиз Я.И. Сооб- щение 4. Морфологические и ультраструктурные изменения печени крыс // Радиобиология. 1991. Т. 31, вып. 5. С. 648 – 653. Пястолова О.А., Трубецкая Е.А. Некоторые морфологические и цитогенетические осо- бенности печени сеголеток Rana arvalis в условиях техногенного ландшафта // Экология. 1989. № 5. С. 57 – 63. Пястолова О.А., Вершинин В.Л. Некоторые цитологические особенности остромордой лягушки на территории Восточно-Уральского радиоактивного следа // Экология. 1999. № 1. С. 26 – 30. Пястолова О.А., Вершинин В.Л., Трубецкая Е. А., Гатиятуллина Э.З. Использование амфибий в биоиндикационных исследованиях территории Восточно-Уральского радиоак- тивного следа // Экология. 1996. № 5. С. 378 – 382. Саркисов Д. С. Регенерация и ее клиническое значение. М., 1970. 175 с. Черданцев В.Г., Ляпков С.М., Черданцева Е.М., Северцов А.С. Методы и результаты анализа экологической устойчивости популяций бурых лягушек при радиационном загряз- нении // Экологические последствия радиационного загрязнения на Южном Урале. М.: Наука, 1993. С. 303 – 307. Черных А.Б., Морозов А.Е., Магда И.Н., Бекенов А.Б., Понявкина А.Г., Мырзабеко- ва Д.М. Внутривидовая изменчивость мышевидных грызунов, населяющих Семипалатин- ский испытательный полигон // Изв. Акад. наук РК. Сер. Биология-Медицина. 1998. Вып. 4. С. 25 – 32. Черных А.Б., Морозов А.Е., Магда И.Н., Бекенов А.Б., Понявкина А.Г., Мырзабеко- ва Д.М. Эколого-эволюционные преобразования в популяциях лесной мыши Семипалатин- ского полигона // Проблемы охраны и устойчивого использования биоразнообразия живот- ного мира Казахстана / Ин-т зоологии и генофонда животных МН-АН РК. Алматы, 1999. С. 48 – 49. Humphrey C., Pittman F.A. Simple methylene blue-azure 2 basic fuchysin stane for embedded tissue sections // Staine Technology. 1974. Vol. 49. P. 9 – 14. Martoja R., Martoja-Pierson U. Initiation aux techniques de l’histologie animali. P., 1967. 345 p. Reynolds E.S. The use of lead citrate at high pH as an electronopaque stain in electron microscopy // J. of Cell Biology. 1963. Vol. 17. P. 208 – 212. Т.Н. Дуйсебаева, Г.В. Федотовских, Н.А. Стрелюхина
HISTOLOGICAL AND ULTRASTRUCTURAL STUDIES OF THE LIVER IN THE LIZARDS FROM THE FORMER SEMIPALATINSK TEST SITE (EASTERN KAZAKHSTAN)
T.N. Dujsebayeva 1, G.V. Fedotovskikh 2, N.А. Strelyukhina 3 1 Institute of Zoology, Ministry of Education and Sciences Аl-Farabi Pr., 93, Almaty, 050060, Kazakhstan 2 Scientific Surgery Center named after Sizganov Zheltoksan Av., 62, Almaty, 050004, Kazakhstan 3 Institute of Experimental Biology Abaya Av., 78, Almaty, 050030, Kazakhstan
The paper presents data on the histology and ultrastructure of the liver of two lizard species, Lacerta agilis and Eremias arguta, inhabiting the territory of the former Semipalatinsk Test Site (Eastern Kazakhstan). An LM study has shown the presence of pathological changes expressed in irregular vascular filling of the liver and venous, local discomplexation of hepatocyte, vacuole and adipose dystrophies and an increase of the local micronecroses where the lymphocyte, resident macrophage and Kupffer cell infiltrations have been registered. An EM investigation has revealed the features of glycogenolysis and decreasing of the lipid embeddings, the phenomenon of hydropical dystrophy and intracellular swelling in the certain part of the hepatocytes, significant hemosiderin accumulations in the liver cells appeared because of intensified catabolism of the phagocyted erythrocytes. The high level of metabolism processes was provided by utilization of the lipid embeddings, the high level of nucleoprotein synthesis and intensification of phosphorylating function of the mitochondria. The functional activity of Kupfer’s hyperplastic cells was directed to the synthesis of pigment granules, the most important role among which belongs to melanin as a strong antioxidant protecting the body against free radicals. Key words: lizards, liver, histology, ultrastructure, nuclear pollution.
Отсюда можно сделать предположение,что если Земля является целью переселения ящероидов,то они постараются не допустить радиоактивного заражения планеты.
Ну, развиваем тему, не стесняемся. Этим праативным ящерицам надо сначала извести радиацией биологических людей, а себе оставить чистую территорию. Раскидываем мозги дальше - какая территория самая свободная от людей и экологически чистая ? Кто правит этой территорией ?
Правильно, я давно говорил, но меня не слушали:
ЯПОНЦЫ - ДЕТИ ЗЕМЛИ, РОССИЯ - СТРАНА ЯЩЕРНЫХ ВЫРОДКОВ, ГЛАВНЫЙ ЯЩЕР РОССИИ - СЕРГЕЙ ИВАНОВ, АЛЕКЗЕС И МУЖИК С КЛИЧКОЙ "ИС" - ЕГО ГЛАШАТАИ И ПРЕСС-СЕКРЕТАРИ, АНТИХРИСТ РОДИТСЯ В РОССИИ !
Хамо, я поймал смысл мессиджа ?
Сообщение отредактировал Индра_Второй - Суббота, 02.04.2011, 10:04
Да уж, граждане товарищи и господа... И террористы у нас все из террариума сбежамши...
Еще раз для Георгиев, гоняющихся за драконом. Анти-крест - ЭТО МАЯК СИДОНИИ - п. МАРС.
Марсиан, не поленитесь, поищите, называют также антами...
И что при аховой ситуации с дебилами, обезьянами и лучевым и волновым оружием, ставшим "не по мозгам" - НЕУДИВИТЕЛЬНО, что ВСЕ оставшиеся в живых маячники участвует в сдерживании. Кому смогли - объяснили, кому не смогли - не серчайте, была возможность понять без религиозной пудры и лапши.
Марсиан, не поленитесь, поищите, называют также антами..
Ant - муравей (англ) Выходит, марсиане у нас Инсектоиды Муравьи наверное не такие страшные, правда, Хамо?
Помните, был такой мультик " Муравей Антц"?
Сюжет(для тех, кто не смотрел) Большой муравейник поделён на две части: там живут муравьи-рабочие и муравьи-солдаты. Рабочие трудятся с утра до ночи, а солдаты воюют с термитами под командованием властного и завистливого генерала Навозника. Так было всегда. И никому, кроме муравья Зэда, любящего пофилософствовать и поворчать (чем он резко отличается от всех муравьев), работающего на строительстве туннеля, не приходит в голову, что все может быть по-другому. Однажды в бар на танцы тайно приходит принцесса Бала и там знакомится с Зэдом. Он влюбляется в красавицу с первого взгляда. Чтобы еще хоть раз увидеть ее, Зэд меняется местами с приятелем-солдатом Уивером и отправляется на военный парад, не ведая, что этим поступком он изменит ход истории…
Можно и с этого начать рассуждения и поиск доказательств. Но вообще-то это тип скафандров, они либо тяжелые десантные, толстые, довольно грубые - вроде драконидных визуализаций, либо летные - тонкопластинчатые, с фасетчатым забралом или без оного, это уже детали.
Примерно вот такие:
Сообщение отредактировал Is - Суббота, 02.04.2011, 17:09
Похож на жука Скарабея. Is, как Вы думаете, какова функциональность накидки, свисающей сзади? Если они своим видом подражали прапрапра... родичам, то это же в чистом виде - сложенные крылья. Вот такие мне снились с пол года назад. Они собирались взорвать какое-то высокое здание. Возили на тележках красные баллоны с с проводами.
Да. Только легкие ткани - были воспроизведением особенностей поля ( плащи, пелерины, сложные воротники). А вот оперение антенн - это уже на шлемах скафандров, по аналогии с антенным оперением кораблей.
А лучевой нанизыватель - для разведки или для точечных ударов?У меня сегодня день почемучки. И очень похоже на собирание картинки из паззлов. Спасибо, Is!
Лучевой нанизыватель нужен для "прокола" пространств со слоистой, нестабильной гравитацией, возможности его зависят от корабельной силовой установки, по ее мощности в основном и шло деление на классы звездолетов, " ходящих по лучу ".
Может использоваться и как оружие " последнего довода".
Гравитационная разведка - другое оборудование, обеспечивающее как раз стабильный режим работы нанизывателя.





































 А потом всякое могло быть,вдруг автор умер,и наследники откопали случайно,или еще какой вариант...
А потом всякое могло быть,вдруг автор умер,и наследники откопали случайно,или еще какой вариант... 
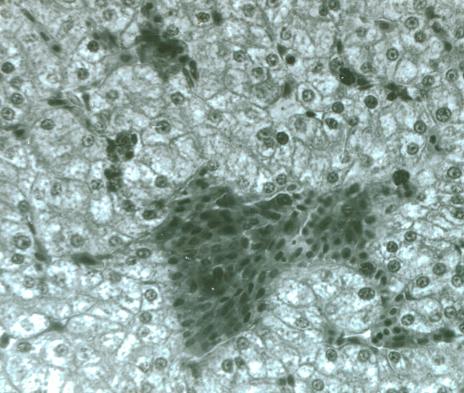
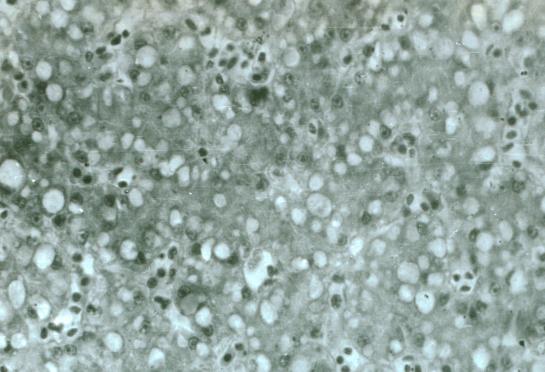

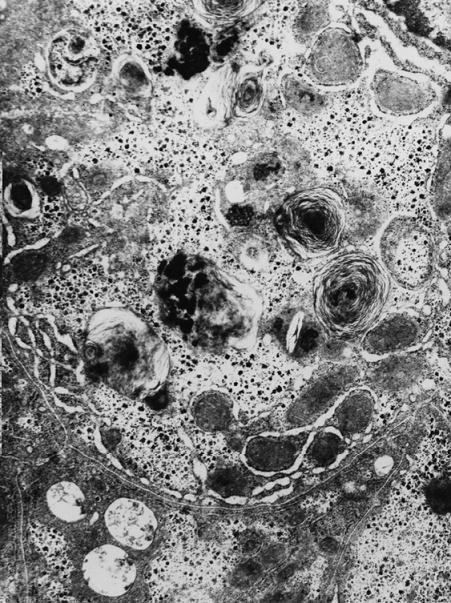
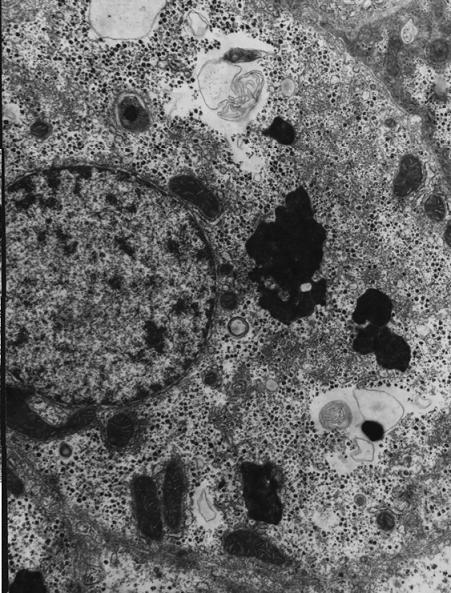
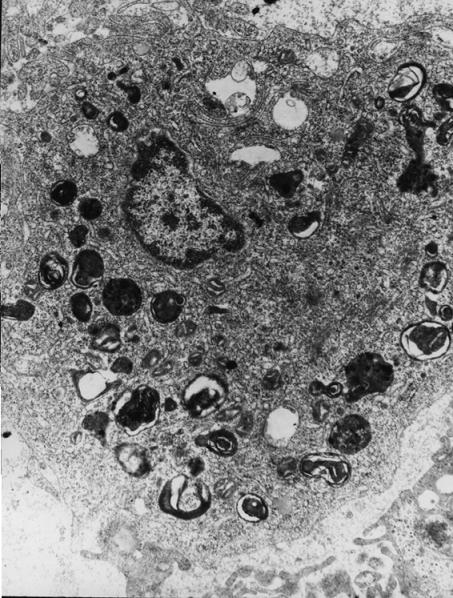











 И очень похоже на собирание картинки из паззлов.
И очень похоже на собирание картинки из паззлов.